
食肉牛龍屬
| 食肉牛龍屬 | |
|---|---|

| |
| 骨架模型,展示於捷克布拉格Chlupáč博物館 | |
| 科學分類 | |
| 界: | 動物界 Animalia |
| 門: | 脊索動物門 Chordata |
| 綱: | 蜥形綱 Sauropsida |
| 總目: | 恐龍總目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亞目: | 獸腳亞目 Theropoda |
| 科: | †阿貝力龍科 Abelisauridae |
| 亞科: | †食肉牛龍亞科 Carnotaurinae |
| 演化支: | †短吻龍類 Brachyrostra |
| 演化支: | †堅背龍類 Furileusauria |
| 族: | †食肉牛龍族 Carnotaurini |
| 屬: | †食肉牛龍屬 Carnotaurus Bonaparte, 1985 |
| 模式種 | |
| †薩氏食肉牛龍 Carnotaurus sastrei Bonaparte, 1985
| |
食肉牛龍(屬名:Carnotaurus,發音:/ˌkɑːrnoʊˈtɔːrəs/,意為「食肉的牛」,有時簡稱為牛龍)是一屬阿貝力龍科的獸腳亞目恐龍,生存於晚白堊世(可能約7200萬至6990萬年前)的南美洲,相當於現在的阿根廷巴塔哥尼亞。唯一的物種是模式種薩氏食肉牛龍(Carnotaurus sastrei)。對其的瞭解來自單一副保存良好的骨骼,而成為南半球所知最詳盡的獸腳類之一。化石於1984年在丘布特省的殖民地組發現。食肉牛龍是阿貝力龍科中的進階型物種,這是一群在晚白堊世的岡瓦納大陸佔據頂級掠食者棲位的中大型肉食性恐龍。牠在科中通常被認為屬於短吻龍類演化支,是一群僅分布於南美洲的短吻部物種。
食肉牛龍是種身體結構輕盈、二足行動的掠食者,經測量估計身長約7.5至9公尺,體重至少1.35噸。以獸腳類來說,食肉牛龍高度特化並且非常獨特。眼睛上方長有厚犄角,這在其他所有獸腳類都未曾觀察到過;縱視非常高深的頭顱長在肌肉發達的粗壯頸部上。此外另一項明顯特徵是極短、已退化成痕跡器官的前肢,搭配長而纖細的後肢。骨骼標本還額外保存了大面積的皮膚印痕,可知牠身上鑲嵌著許多小塊(直徑約5毫米)、不重疊的鱗片,身體兩側長著許多大型、不規則排列的圓形骨質突起物(皮內成骨),沒有羽毛的跡象。
獨特的犄角和肌肉發達的頸部可能是使用於物種內打鬥。根據不同的研究,互相競爭的個體可能在打鬥時使用頭部來撞擊,以頭骨上側緩慢互推、或將角作為緩衝正面撞擊對方。尚不清楚食肉牛龍的覓食習性:某些研究認為牠有能力撂倒非常大型的蜥腳類恐龍,其他研究則認為牠主要獵捕相對小型的獵物。腦腔結構暗示有著非常發達的嗅覺,而聽覺和視覺較不發達。食肉牛龍可能擅於奔跑,並可能是行動最迅速的大型獸腳類之一。
發現
[編輯]

食肉牛龍的唯一一具標本(正模標本MACN-CH 894)是由阿根廷古生物學家何塞·波拿巴所率領的考察團隊於1984年挖掘出土。[A]同時還發現了擁有特殊棘刺的蜥腳類阿馬加龍。[3]此次挖掘活動屬於「南美洲侏羅紀與白堊紀陸生脊椎動物」計畫的第八次考察,該計畫始於1976年,並由國家地理學會贊助。[3][B]骨骼保存良好且關節相連,只缺乏尾部後段三分之二、大部分的下肢骨、後腳已被風化分解。[C][5]根據腦殼癒合的縫線來判斷,這具骨骼屬於一個成年個體。[6]牠被發現時倒於右側,呈現典型的死亡姿勢,頸部和背部沿軀幹向後彎曲。[7]不尋常的是,有大片的皮膚印痕保存下來。[D]有鑑於這些印痕的重要性,對原來的挖掘地點展開了第二次的考察,過程中又找到了更多皮膚印痕。[7]頭骨在化石化過程中變形了,左側吻部骨頭相對右側向前位移,鼻骨被向上擠壓,前上頜骨向後推向鼻骨之上。變形也讓上頜上彎的程度過度加大。[E]吻部受到變形的影響比頭骨後部來得嚴重,可能是因為後者較為堅固。從頂部或底部視角來看,上頜的U形相對於下頜的U形較不明顯,導致兩者明顯不相契合。這樣的不相契合是由側向作用的變形所造成,上頜被影響而下頜沒有,可能是因為後者關節柔韌性較好。[1]
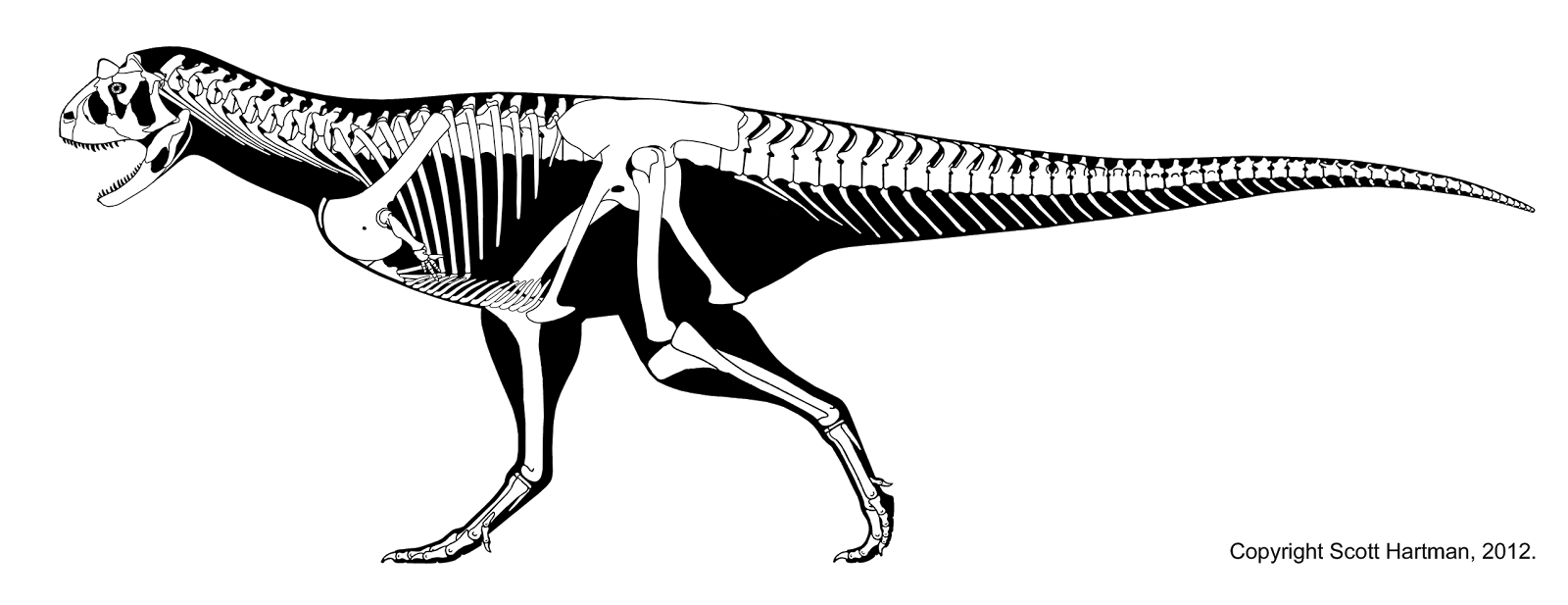
化石採集於阿根廷巴塔哥尼亞丘布特省特爾森縣棕坡(Bajada Moreno)附近的薩斯特雷農場(Pocho Sastre)。[5]因為埋在十分堅硬的大型赤鐵礦結核岩塊中,使得清修過程複雜並進展緩慢。[9][5]1985年,波拿巴發表了一篇筆記,介紹了新屬新種薩氏食肉牛龍,並簡略描述了頭骨與下頜。[5]屬名源自拉丁語的carno/carnis(肉)和taurus(公牛),意即「食肉的牛」,象徵著形狀像牛的犄角。[10]種名sastrei紀念化石發現地牧場的擁有者安赫爾·薩斯特雷(Angel Sastre)。[11]1990年,波拿巴、費爾南多·諾瓦斯和魯道夫·科里亞發表了一份關於整副骨骼的全面性描述論文。[4]食肉牛龍是繼阿貝力龍後,阿貝力龍科第二個發現的物種。[12]多年來牠一直是該科、同時也是南半球獸腳類中,了解最詳盡的物種。[13][14]直到進入21世紀相繼敘述了更多保存良好的阿貝力龍科(如奧卡龍、瑪君龍、蠍獵龍)之後,才使研究人員能夠針對食肉牛龍解剖學的特定方面進行重新檢驗與評估。[F]正模標本目前保存於阿根廷自然歷史博物館;[G]複製重建骨架模型在那裡以及世界各地的博物館都有展設。[15]雕刻家史蒂芬與希薇亞·澤卡斯(Stephen and Sylvia Czerkas)製作的一個等比例復原模型以前曾在洛杉磯縣自然歷史博物館展示過。這個雕塑作品是博物館於1980年代中期訂製的,可能是首具高準確性皮膚還原度的獸腳類復原模型。[7][16]
敘述
[編輯].png)
體型
[編輯]
食肉牛龍是種大型、但結構輕盈的掠食者。[17]唯一已知的標本身長約7.5至9公尺,[H][I][19]成為體型最大的阿貝力龍科之一。[J][K][19]爆誕龍及可能的阿貝力龍(化石都不完整)體型可能相近或更大。[L][M][N]一份2016年的研究指出,只有身長8.9公尺的密林龍,長於食肉牛龍的估計值7.8公尺。[21]頭部高2.9公尺。[22]體重在不同的研究中使用不同方法測量曾被估計為1350公斤[O]、1500公斤[P]、2000公斤[19]、2100公斤[Q]、2500公斤[22]、以及1306至1743公斤[25]。由頭骨、脊椎、前肢的特徵可以發現,食肉牛龍是高度特化的獸腳類。[R]另一方面,坐骨和後肢則相對原始,類似更基礎的角鼻龍。坐骨和後肢都長而纖細:其中左股骨測量長103公分,但平均直徑只有11公分。[S]
頭骨
[編輯]
頭骨測量長59.6公分,與其他大型肉食恐龍相比,食肉牛龍的頭部比例上較小而高度深。[T][U]吻部略寬,不像基礎獸腳類(如角鼻龍)那樣尖細,下巴向上彎曲。[26]一對明顯的角從眼睛上方斜向突出,由額骨構成,[V]厚、呈圓錐形,內部結實,垂直剖面略為扁平,測量長15公分。[6][1]波拿巴於1990年提出這對角可能構成更長的角質鞘骨核。[W]摩里西奧·瑟隆尼(Mauricio Cerroni)和同事於2020年認同角覆蓋著角質鞘,但反駁道這些角質鞘不會長於骨核本身。[1]

如同其他恐龍,頭骨兩側各有六個主要孔洞。最前面的鼻孔半矩形朝向側前,但側面看不像其他角鼻龍類那樣傾斜;僅由鼻骨和前上頜骨構成,而一些近緣物種的上頜骨也有參與構成。在鼻孔和眼窩之間是眶前孔。角鼻龍的眶前孔高度大於長度,但相關物種如蠍獵龍、瑪君龍的是長度大於高度。眶前孔由較大的凹陷眶骨窩所包圍,前部由上頜骨凹部、後部由淚骨構成。如同所有阿貝力龍科,食肉牛龍的這個凹陷較小。眶骨窩前下角有個較小的孔洞前上頜孔,通往上頜骨內的空腔。[1]眼睛位於鑰匙孔形的眼眶上部。[X]上部比例小、呈圓形,以前突的眶後骨與眼眶下部分隔開來。[1]眼睛略轉向前,可能具有一定程度的雙眼視覺。[Y]如同所有阿貝力龍科,額骨(位於雙眼之間的頭蓋骨)不包含在眼眶內。眼眶後面有兩個孔洞:側面是下顳孔、上面是上顳孔。下顳孔高、短、呈腎形;上顳孔短、呈方形。下頜有另一個孔洞為下頜孔,食肉牛龍的相對較大。[1]
上頜兩側的牙齒數量分別為:前上頜骨4顆、上頜骨12顆;[Z]下頜兩側齒骨各有15顆牙齒。[AA][1]牙齒細長,和其他阿貝力龍科很短的牙齒相反。[26]但瑟隆尼等人(2020)的頭骨敘述指出,所有長出的牙齒在挖掘過程中都受到嚴重損壞,後來才用石膏進行重建;波拿巴(1990)只提到一些下頜牙齒變成碎片。[1][AB]因此關於牙齒形狀的可靠資訊僅限於汰換的牙齒和依然被下頜包覆的齒根,並可透過CT掃描成像來研究。[1]汰換的牙齒有低而平坦的齒冠,排列緊密,前傾約45度。[1]波拿巴(1990)指出下頜淺、結構脆弱,齒骨(下頜最前面的骨頭)僅透過兩個接觸點與後面的下頜骨連接;這與外觀厚實的頭骨形成對比。[9][AC]瑟隆尼等人(2020)卻在齒骨和後部下頜骨之間發現多個鬆散的連接,這使得關節結構可動性高,但不一定脆弱。[1]齒骨底緣凸起,而瑪君龍的則為平坦。[1]
下頜發現在生前位置有成骨作用的舌骨,這條纖細的骨頭附著舌頭等幾條肌肉,這在恐龍中很少見,牠們的舌頭通常屬於軟骨,未與其他骨頭相連,因此很容易遺失。[AD][27][1]食肉牛龍保存了三個舌骨:一對彎曲棒狀的角枝(ceratobranchials)與一個梯形基舌骨(basihyal)關節連接。食肉牛龍是非鳥類獸腳類中唯一已知有基舌骨的。[1]頭骨後面有幾個發育良好的中空腔室圍繞著腦殼,如同所有阿貝力龍科。有兩個獨立的腔室系統:副鼓膜(paratympanic)系統連接中耳空腔、以及頸部起囊膨脹產生的空腔。[25]
頭骨可觀察到數個自衍徵:一對犄角;非常短而高度深的頭骨;上頜骨在前上頜骨孔上方嵌入,本應嵌於眶上氣竇(吻部的空腔);運輸淚液的鼻淚管經過一條未知功能的導管從淚骨內面通出。其他可能的自衍徵有方骨深長中空的洞以及齶翼骨上的加長凹陷。[1]
脊椎
[編輯]
脊柱由10節頸椎、12節胸椎、6節癒合的薦椎[AE]、以及數量未知的尾椎所構成。[4]頸部幾乎筆直,不像其他獸腳類普遍的S形曲線,並且向基部異常地寬。[28]頸柱頂部有兩條向上地骨質突起,稱為上骺,在頸椎頂部構成一道光滑的溝槽。這些突起是整個頸柱的最高點,豎立於異常低矮的棘突之上。[4][27]上骺可能附著著強壯的頸部肌肉組織。[AF]類似的兩條結構也可在尾部觀察到,由高度特化的尾肋所構成,前面看來呈V形且向上突出,內側構成了前部尾椎光滑、平坦的頂面。每個尾肋末端有前突的鉤形擴張,與前續脊椎的尾肋相連。[27][29]
頸椎的結構類似現代鳥類,可以做出更快、更準確的動作。薦椎數量相當多,共有8節,可以承受碰撞和衝擊。[4]
前肢
[編輯]
食肉牛龍的前肢極為短小,甚至比任何大型獸腳類(包括暴龍)都還要短。[AG]前臂只有上臂長度的四分之一。手部沒有腕骨,因此掌骨直接與前臂關節連接。[30]手部有四根基本的手指,雖然其中只有中間兩根以指骨收尾,而第四指僅由一塊夾板狀的掌骨構成,可能形成一種外部「骨刺」。手指本身癒合無法動作,可能沒有爪子。[5][4]食肉牛龍與其他阿貝力龍科的不同點在於:前肢比例較短但更健壯、第四指夾板狀的掌骨是手部最長的骨頭。[31]2009年的一項研究指出,手臂在阿貝力龍科已經退化成痕跡器官,上頭的神經傳導纖維已經減少至相當於現代的鴯鶓或奇異鳥(都有前肢痕跡器官)的程度。[32]

前後肢比例的比較:食肉牛龍的前肢/後肢長度比例為14%、艾伯塔龍的前肢/後肢比為20%、似鴕龍為46%、始祖鳥為96%。肱骨、橈骨、掌骨長度的比較:食肉牛龍分別為28.5公分、7.3公分、3.7公分;艾伯塔龍為32.4公分、15.6公分、9.8公分;懼龍為35.7公分、17.1公分、12公分。由此顯示出,食肉牛龍與暴龍科的前肢比例非常類似,似乎是平行演化的結果。肱骨/橈骨/掌骨百分比例的比較:食肉牛龍為72:19:9,而其他前肢小的獸腳類則為55:27:19左右。[33]大多數獸腳類前肢掌心方向為彼此合掌,而食肉牛龍前肢掌心略朝後上方,目前仍不清楚功用為何。[34]
後肢
[編輯]食肉牛龍的後肢並沒有完整地保存下來,但可以推測其後腿長而強壯,脛部結實,踝關節高,與腳趾間的部份延長、纖細,顯示食肉牛龍是種擅於奔跑的動物。[4]
皮膚
[編輯]
食肉牛龍是第一種發現大量化石皮膚印痕的獸腳類。[7]這些印痕發現於身體的右側下方,來自不同的身體部位,包括下頜、[7]前面的頸部、胸帶、以及肋腔等。[AH]最大塊的來自尾部前面。[AI]起初頭骨右側也覆有大塊皮膚印痕,但清修時沒能及時辨認出來而意外地破壞掉了。[7]但還是可以從頭骨幾個表面紋理推測出可能的表皮狀況。吻部側面及正面發現了一個由凹槽、凹洞及小孔洞的丘狀表面,暗示著鱗片般的表皮,可能像現代鱷魚般的平坦鱗片。吻部頂端裝飾著許多小孔和棘刺,可能與表面覆蓋角質有關,類似的構造也在瑪君龍頭上觀察到,但阿貝力龍和皺褶龍則缺乏。可以確定的是,一圈大塊的鱗片包圍著眼睛,根據淚骨及眶後骨上縱溝及丘狀表面所示。[1][AJ]
皮膚由直徑約5毫米的不重疊多邊形鱗片鑲嵌構成,鑲嵌之間由細的平行溝槽隔開。[AK]身體各部位的鱗片比例大同小異,除了頭部之外,顯然反映出不同、不規則的鱗片形狀。[AL][15]沒有任何羽毛的跡象。[7]
沿著頸部、背部與尾部側面的皮膚上,有許多不規則排列的圓錐形皮內成骨。每個突起都呈低矮稜脊,部分直徑測量約4至5公分。彼此間隔8至10公分,並朝身體上方逐漸變大。這使食肉牛龍的外表凹凸不平。這些隆起可能是密集的鱗甲,類似在鴨嘴龍類皮膚上觀察到沿身體中線排列的柔軟皺褶。[5][AM][7]史蒂芬·澤卡斯(Stephen Czerkas)於1997年提出,這樣的結構可能在物種內或種間打鬥時保護身體側面,並認為在現代鬣蜥頸部可觀察到類似結構,在打鬥時提供有限的防護。[7]
分類
[編輯]食肉牛龍是被研究最透徹的阿貝力龍科之一,牠們是一群僅分布於南方岡瓦那大陸的大型獸腳類,並在白堊紀晚期成為當地的優勢掠食動物,取代鯊齒龍科在白堊紀早期的生態棲位,而同時北半球佔據對應棲位則是暴龍科。[17]該科物種演化出許多顯著特徵,包含變短的頭骨與前肢、以及特殊的頸椎和尾椎結構,後者在食肉牛龍中發展得又比其他阿貝力龍科還要明顯。[AN][AO][29]
雖然阿貝力龍科內的親緣關係備受爭議,系統發生學研究普遍顯示出食肉牛龍是科內最衍化的進階型物種。[AP]牠的最近緣物種可能是奧卡龍[35][36][37][38]或瑪君龍[39][40][41]。但2008年馬修·卡拉諾(Matthew T. Carrano)和史考特·桑普森的角鼻龍類系統發生學研究得出相反的結果,認為食肉牛龍與前兩者都不近緣,而是與肌肉龍互成姊妹群。[AQ]
1998年,芝加哥大學的保羅·塞里諾提出食肉牛龍亞科(Carnotaurinae),定義為:除了在許多研究被認為是基礎物種的阿貝力龍以外,包含所有的進階型阿貝力龍科物種。[42][43]但2008年的研究認為阿貝力龍也是進階型物種。[AR]

2002年,科里亞等人在新敘述的奧卡龍身上找到數個和食肉牛龍相同的共有衍徵,尤其是尾部近段[註 1]與中段具有下椎弓突-下椎弓凹關節,且頸椎上突的顱端[註 2](與尾端[註 3][35])具有突起,這兩點都僅見於這兩種恐龍,有別於其他的阿貝力龍科,因此他們將食肉牛龍和奧卡龍劃分進新的演化支:食肉牛龍族(Carnotaurini),代表兩者為最近緣物種。[36][44]然而,食肉牛龍亞科和食肉牛龍族並不被學界廣泛接受。
以下演化樹取自塞里諾和同事(2004)關於阿貝力龍類地理分布與系統發生學的研究,參考科里亞等人(2002)的數據:[39]
| 阿貝力龍科 Abelisauridae |
| ||||||||||||||||||||||||||||||||||||

2009年,胡安·卡納雷(Juan Canale)和同事的系統發生學研究,將南美洲的食肉牛龍亞科加上肌肉龍,建立成一個食肉牛龍亞科內的次演化支短吻龍類(Brachyrostra),定義為:阿貝力龍科中,與食肉牛龍的親緣關係比與瑪君龍還要更近的物種;這項分類被許多後來的研究所採納。[35][38][45]其演化樹如下所示:[35]
| 食肉牛龍亞科 Carnotaurinae |
| ||||||||||||||||||||||||||||||
2013年,提爾黎·托爾托薩(Thierry Tortosa)等人提出新版本的阿貝力龍科演化樹。主要可區分為三大類:基礎型的隱面龍(非洲)、皺褶龍(非洲)、怪踝龍(南美洲)、膝龍(歐洲)、塔哈斯克龍(歐洲);瑪君龍亞科有印度的勝王龍、拉休里龍、馬達加斯加的瑪君龍以及法國的阿克獵龍;短吻龍類全部分布於南美洲,其中肌肉龍、爆誕龍、蠍獵龍為基群,而阿貝力龍、奧卡龍、基爾梅龍、密林龍及食肉牛龍一起構成最進階的演化支。[46]
| 阿貝力龍科 Abelisauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
2015年,在雷納多·費里皮(Leonardo S. Filippi)等人敘述道獵龍的研究中,建立了新的演化支堅背龍類(Furileusauria),以包含較進階的短吻龍類,其中食肉牛龍成為食肉牛龍亞科的唯一物種,而奧卡龍和阿貝力龍則位於阿貝力龍亞科(Abelisaurinae),於是先前的食肉牛龍族就變成單系群而顯得多餘了。[47]
| 阿貝力龍科 Abelisauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
古生物學
[編輯]額角的功能
[編輯]
目前已發現許多獸腳類恐龍的頭部具有骨質冠飾:有些是縱向的冠飾,例如合踝龍、雙脊龍與冰脊龍;有些則是鼻部的角狀物或隆起,例如原角鼻龍與角鼻龍;食肉牛龍的兩個角不位於鼻部,而位於額頭上,是唯一已知額骨上有一對角的二足肉食性動物。[48]關於這對角的功能並不是很清楚,有些詮釋成用在種間/內打鬥行為或殺死獵物,有些則認為用於求偶展示或種內辨識也是有可能的。[1]
葛瑞格利·保羅(1988)提出角是緩衝用的武器,其小弧度能使打鬥時造成眼睛受傷的可能降到最低。[9]格拉多·馬澤塔(Gerardo Mazzetta)等人(1998)認為食肉牛龍角的功能類似於羊的犄角。他們計算頸部肌肉夠強壯,足以吸收兩個個體以5.7公尺/秒的速度正面衝擊的力道。[23]諾瓦斯(2009)解釋了許多種用來適應頭部撞擊的骨骼特徵。[AS]他認為短頭骨可減少轉動慣量使頭部運動速度加快,而頸部肌肉能夠承受強烈的頭部衝擊。他亦指出脊柱剛性和強度增強可能是發展用來承受頭部與頸部的衝擊。[AT]
其他研究則認為互相競爭的食肉牛龍無法快速頭部撞擊,而是以頭頂緩慢地互相推擠。[48][49]馬澤塔等人(2009)表示角可能是用來分散壓力不讓腦部受傷的一種構造,並根據角的上側平坦、頭頂骨大幅癒合、以及頭骨無法承受快速撞擊來支持此理論。[48]拉斐爾·德庫特(Rafael Delcourt)(2018)認為這個角可能用來像現代海鬣蜥一樣的慢速頭部推撞,或是像長頸鹿一樣打擊對手頸部及身體側面。[38]後者的可能性曾在先前2011年相關物種瑪君龍的研討會論文中提出過。[50]
馬澤塔等人(1998)提出角可能用來弄傷或殺死小型獵物。雖然角核較鈍,但若加上角質覆蓋物其形狀可能近似現代的牛角。然而,這將成為把犄角當作狩獵武器的唯一例子。[23]
頜部功能與咬合力
[編輯]
與類似體型的獸腳類相比,食肉牛龍的頭部小、短而寬,如同其他阿貝力龍科,其嘴部大量小型鋸齒狀牙齒適合撕咬獵物、而短吻部具有更大的咬合力。雖然食肉牛龍頭部較為短小,但研究顯示牠們的頭部有良好的力學結構。馬澤塔等人於1998年、2004年及2009年的研究對食肉牛龍頜部進行分析,認為牠有能力快速的咬合,但咬合力量不強勁。[23][24][48]當捕捉小型獵物時,快咬會比強咬來得重要,正如同現代鱷魚研究發現的情況。[48]研究人員還指出頭骨中、尤其是下頜充滿高度靈活性(頭部可動性),某種程度上近似現代的蛇類。頜部的彈性使食肉牛龍能夠將小型獵物整隻吞下。此外下頜由關節相連而能夠上下移動。下壓時牙齒向前突出,從而刺入小型獵物;當牙齒向上收合時,向後突出的牙齒能夠固定獵物防止逃逸。[23]他們還發現頭骨能夠承受撕扯大型獵物產生的壓力。[48]食肉牛龍可能主要獵捕小型動物,但也有能力掠食大型恐龍。[48]愛蜜莉·瑞菲德(Emily Rayfield)(2001)使用有限元分析研究獸腳類頭骨,以電腦斷層掃描重建出頭骨與下頜的數位模型,發現食肉牛龍的頂骨與額骨之間緊密地癒合,使頭骨可以承受相當高的壓力。位於額頂的兩個角,使頭頂的形狀略顯寬、平,可以承受碰撞。這個有限元分析模型將頭部肌肉分解成四個聯結的部份來計算其咬合力,包含:顳肌、翼肌、下頜收肌(Adductor mandibularis muscle)、下頜內肌(Intramandibular muscle)。[51]保羅·巴雷特(Paul M. Barrett)與瑞菲德(2006)研究獸腳類咬合時的肌肉力學和上下頜閉合速度,發現食肉牛龍有較快的咬合速度;而角鼻龍與暴龍的咬合速度較慢但咬合力更大。[23][52]馬澤塔(2004)估計食肉牛龍的咬合力最大值為3341牛頓(嘴部後段),與其他體型接近的獸腳類相比較弱,因此可能無法在獵食過程中咬碎大型獵物的骨頭。當食肉牛龍咬合時,產生的咬合力小,而頭骨結構能承受巨大壓力,頭部肌肉大幅地減少來自上頜的應力與形變,連結上下頜的頜部關節可以減低咬合時的衝擊。嘴部開闔幅度大、咬合速度快、牙齒小呈鋸齒狀,並有聯結頭部與頸部的肌肉群,顯示牠們可能採取撕咬的方式攻擊獵物。[24]
以上詮釋受到法蘭索瓦·席瑞恩(François Therrien)等人(2005)的質疑,他們發現食肉牛龍的咬合力,是現生四足類中咬合最強的美國短吻鱷的兩倍。同時也指出,與現代科摩多巨蜥作比較,下頜彈韌度朝尖端線性減低,代表頜部不適合高精準度的捕食小獵物,而適合創傷大型獵物使之虛弱。因此根據這個研究的推測,食肉牛龍必定主要掠食大型動物,可能以伏擊的方式。[53]瑟羅尼等人(2020)反駁道下頜彈性受限,而頭蓋骨變厚和許多頭骨關節骨化代表頭骨沒有或很少可動性。[1]
羅伯特·巴克(1998)認為食肉牛龍主要掠食非常大型的獵物,尤其是蜥腳類。他指出,在侏羅紀晚期的北美洲,生存着許多蜥腳類恐龍,而當時的大型掠食者異特龍出現許多適應性特徵,包括吻部縮短、牙齒相對較小、頭骨後部(枕骨)增強;在白堊紀的南美洲,蜥腳類同樣十分繁盛,使食肉牛龍發生趨同演化。兩者相似的適應特徵都是以上頜作為鋸棍來劃傷、並反覆攻擊、削弱蜥腳類。[54]
運動
[編輯]

馬澤塔等人(1998,1999)推測食肉牛龍是敏捷的奔跑者,並反駁股骨適應來承受跑步時的高度彎曲動作。動物腿部承受這種壓力會限制其最高速度。食肉牛龍的奔跑適應優於人類,但不及鴕鳥。[AU][55]科學家計算食肉牛龍最高速度每小時可達48至56公里。[56]
在恐龍中,最重要的運動肌肉位於尾部。這條肌肉稱為尾股肌,附著於大腿骨上明顯的嵴第四轉子,收縮時將大腿骨向後拉。史考特·佩森斯(Scott Persons)和菲力·柯里(2011)指出,在食肉牛龍的尾椎中,尾肋並非水平突出(T字形),而是斜靠脊椎的垂直軸成V字形。這將提供比任何其他獸腳類都還要大的額外尾股肌空間,計算每條腿的肌肉質量介於111至137公斤。因此食肉牛龍可能是速度最快的大型獸腳類之一。[29]雖然尾股肌加大,位於尾肋之上的軸上肌比例較小。這些肌肉稱為最長肌和棘肌,負責尾部的動作與穩定性。在這些肌肉減少卻又要同時維持尾部的穩定性,尾肋帶有前突將脊椎彼此間及與骨盆相互鎖緊,使尾部變得僵硬。於是用力彎曲的能力減弱,因為臀部和尾部必須同時彎曲,這點與其他獸腳類不同。[29]
腦部與感官
[編輯]瑟隆尼和亞莉安娜·寶琳娜-卡拉巴賈(Ariana Paulina-Carabajal)(2019)使用CT掃描來研究包含腦部的顱腔。顱腔容量為168.8立方公分,而腦部僅佔一小部分的空間。作者估計了兩種不同尺寸的腦部,分別為佔顱腔的50%和37%。此結果在爬蟲類腦化指數中大於關聯物種瑪君龍、但小於暴龍科。製造激素的松果體可能比其他阿貝力龍的還小,由硬腦膜擴張程度低(前腦頂部的一個空間,認為松果體位於其中)來判斷。[25]
載有嗅覺的嗅球大,而負責視覺的上丘相對較小。這代表嗅覺優於視覺,而現生鳥類的情況則相反。嗅管和嗅球下彎,只有印度龍有相同的特徵。如同作者所假設,這樣的下彎和大尺寸嗅球可能代表著食肉牛龍比起視覺更依賴的是嗅覺。被認為與穩定凝視有關/眼睛和身體之間的協調的腦葉-小腦鬈,在食肉牛龍和其他阿貝力龍科都很大。這可能代表這些動物經常將頭部和身體作快動作。聽覺可能在食肉牛龍和其他阿貝力龍科發育程度弱,如內耳的聽壺所示。估計聽力範圍低於3千赫茲。[25]
古生態學
[編輯]
食肉牛龍化石最初被認為發現於赤山組的上部岩層,年代約1億年前(阿爾比階或森諾曼階)。[5][AV]但這個地層的年代被認為不是很精確。後來他們才意識到其實是屬於更年輕的殖民地組,估計年代大致介於坎潘階與馬斯垂克階,約8360萬至6600萬年前。[13][57][1]諾瓦斯在他2009年的書籍中定出了更精確的年代,為7200萬至6990萬年前的馬斯垂克階早期。[AW]食肉牛龍因而成為已知年代最晚的南美洲阿貝力龍科。[29]在晚白堊世期間,南美洲大陸已經從非洲和北美分裂孤立出來。[58]
殖民地組暴露於北巴塔哥尼亞地塊南坡上。包括食肉牛龍的大部分脊椎動物化石都來自地層中相段。當地被認為過去可能是個河口灣、泥灘或海岸平原的沉積環境,,具有沙灘,位於河流出海口,並可能為乾濕季兼具的季節性氣候。[59]這裡有發現軟體動物(例如雙殼類)的化石,應該是卡瓦斯海(Kawas sea)(一個50萬平方公里的遠古海洋)海侵的結果。[59]當地最常見的脊椎動物是角齒魚科的肺魚、烏龜、鱷魚、蛇頸龍類、恐龍、蜥蜴、蛇以及哺乳類。[60]蛇類有蚺科和巨蛇科,例如阿根廷阿拉米巨蛇。[61]烏龜以至少五個物種為代表,四種側頸龜亞目蛇頸龜科與一種曲頸龜亞目捲角龜科。[62]海生爬蟲有蛇頸龍目雙臼椎龍科的槽鱷龍。[62]哺乳類有瑞吉獸(首個在南美洲發現的柱齒獸類)[59],以及可能屬於多瘤齒獸目的阿根廷旅獸。[63]2011年新發現一種反鳥類。[64]近年來在殖民地組發現了許多種水生植物,包含蕨類、裸子植物、被子植物,[65][66]如Paleoazolla與Regnellidium。[67]還發現了南美洲已知最早的蓮科果實化石。[68]
大眾文化
[編輯]從1990年代中期開始,食肉牛龍偶爾出現在大眾媒體之中。食肉牛龍最早出現在麥可·克萊頓1995年的小說《侏羅紀公園2:失落的世界》,故事中的食肉牛龍被作者添加了類似變色龍或章魚依所處環境改變外表顏色的能力,但這僅止於想像。[69]1997年的同名改編電影中,食肉牛龍並未被採用,但有出現在數個周邊商品,如模型玩具、電腦遊戲。一直到2018年的新系列《侏羅紀世界:殞落國度》中,食肉牛龍才終於登場。[70]
在迪士尼2000年的動畫電影《恐龍》中,食肉牛龍作為主要的反派。但片中的食肉牛龍體型被誇大成接近暴龍,現實中卻比該片的主角禽龍還小。[71]
美國科幻電視劇《史前新紀元》中食肉牛龍作為白堊紀晚期的頂級掠食者登場,會主動捕食移居至史前新世界殖民地的人類,也會獵食其他恐龍。劇中牛龍體型比現實巨大,而且手臂更長,具有類似羽毛的構造。
2022紀錄片《史前地球》中也有登場,片中雄性食肉牛龍利用顏色鮮豔的前肢吸引異性。
參見
[編輯]備註
[編輯]參考來源
[編輯]- ^ p. 276 in Novas (2009)[2]
- ^ p. 2 in Bonaparte (1990)[4]
- ^ p. 2 in Bonaparte (1990)[4]
- ^ p. 2 in Bonaparte (1990)[4]
- ^ p. 191 in Carrano and Sampson (2008)[8]
- ^ p. 191 in Carrano and Sampson (2008)[8]
- ^ p. 3 in Bonaparte (1990)[4]
- ^ p. 38 in Bonaparte (1990)[4]
- ^ p. 162 in Juárez Valieri et al. (2010)[18]
- ^ p. 191 in Carrano and Sampson (2008)[8]
- ^ p. 162 in Juárez Valieri et al. (2010)[18]
- ^ p. 163 in Juárez Valieri et al. (2010)[18]
- ^ p. 556 in Calvo et al. (2004)[20]
- ^ p. 191 in Carrano and Sampson (2008)[8]
- ^ p. 30 in Bonaparte (1990)[4]
- ^ p. 187 in Mazzetta et al. (1998)[23]
- ^ p. 79 in Mazzetta et al. (2004)[24]
- ^ p. 276 in Novas (2009)[2]
- ^ p. 28–32 in Bonaparte (1990)[4]
- ^ p. 8 in Bonaparte (1990)[4]
- ^ p. 191 in Carrano and Sampson (2008)[8]
- ^ p. 4–5 in Bonaparte (1990)[4]
- ^ p. 5 in Bonaparte (1990)[4]
- ^ p. 3 in Bonaparte (1990)[4]
- ^ p. 191 in Mazzetta et al. (1998)[23]
- ^ p. 255 in Novas (2009)[2]
- ^ p. 6 in Bonaparte (1990)[4]
- ^ p. 6 in Bonaparte (1990)[4]
- ^ p. 6 in Bonaparte (1990)[4]
- ^ p. 6 in Bonaparte (1990)[4]
- ^ p. 191 in Carrano and Sampson (2008)[8]
- ^ pp. 257 in Novas (2009)[2]
- ^ p. 1276 in Ruiz et al. (2011)[30]
- ^ p. 32 in Bonaparte (1990)[4]
- ^ p. 32 in Bonaparte (1990)[4]
- ^ p. 3 in Bonaparte (1990)[4]
- ^ pp. 264–299 in Novas (2009)[2]
- ^ pp. 264–299 in Novas (2009)[2]
- ^ p. 32 in Bonaparte (1990)[4]
- ^ p. 276–277 in Novas (2009)[2]
- ^ pp. 256–261 in Novas (2009)[2]
- ^ pp. 188–189 and 202 in Carrano and Sampson (2008)[8]
- ^ p. 202 in Carrano and Sampson (2008)[8]
- ^ p. 202 in Carrano and Sampson (2008)[8]
- ^ pp. 259–261 in Novas (2009)[2]
- ^ pp. 260–261 in Novas (2009)[2]
- ^ p. 186 and 190 in Mazzetta et al. (1998)[23]
- ^ p. 3 in Bonaparte (1990)[4]
- ^ p. 276 in Novas (2009)[2]
- ^ 1.00 1.01 1.02 1.03 1.04 1.05 1.06 1.07 1.08 1.09 1.10 1.11 1.12 1.13 1.14 1.15 1.16 1.17 1.18 1.19 Cerroni, M. A.; Canale, J. I.; Novas, F. E. The skull of Carnotaurus sastrei Bonaparte 1985 revisited: insights from craniofacial bones, palate and lower jaw. Historical Biology. 2020: 1–42. doi:10.1080/08912963.2020.1802445.
- ^ 2.00 2.01 2.02 2.03 2.04 2.05 2.06 2.07 2.08 2.09 2.10 Novas, Fernando E. The age of dinosaurs in South America. Bloomington: Indiana University Press. 2009. ISBN 978-0-253-35289-7.
- ^ 3.0 3.1 Salgado, Leonardo; Bonaparte, José F. Un nuevo sauropodo Dicraeosauridae, Amargasaurus cazaui gen. et sp. nov., de la Formacion La Amarga, Neocomiano de la Provincia del Neuquén, Argentina. Ameghiniana. 1991, 28 (3–4): 334 (西班牙語).
- ^ 4.00 4.01 4.02 4.03 4.04 4.05 4.06 4.07 4.08 4.09 4.10 4.11 4.12 4.13 4.14 4.15 4.16 4.17 4.18 4.19 4.20 4.21 4.22 4.23 4.24 4.25 Bonaparte, José F.; Novas, Fernando E.; Coria, Rodolfo A. Carnotaurus sastrei Bonaparte, the horned, lightly built carnosaur from the Middle Cretaceous of Patagonia (PDF). Contributions in Science. 1990, 416: 1–41. (原始內容 (PDF)存檔於July 21, 2010). (頁面存檔備份,存於網際網路檔案館)
- ^ 5.0 5.1 5.2 5.3 5.4 5.5 5.6 Bonaparte, José F. A horned Cretaceous carnosaur from Patagonia. National Geographic Research. 1985, 1 (1): 149–151.
- ^ 6.0 6.1 Paulina Carabajal, Ariana. The braincase anatomy of Carnotaurus sastrei (Theropoda: Abelisauridae) from the Upper Cretaceous of Patagonia. Journal of Vertebrate Paleontology. 2011, 31 (2): 378–386. doi:10.1080/02724634.2011.550354.
- ^ 7.0 7.1 7.2 7.3 7.4 7.5 7.6 7.7 7.8 Czerkas, Stephen A.; Czerkas, Sylvia J. The Integument and Life Restoration of Carnotaurus. Wolberg, D. I.; Stump, E.; Rosenberg, G. D. (編). Dinofest International. Academy of Natural Sciences, Philadelphia. 1997: 155–158.
- ^ 8.0 8.1 8.2 8.3 8.4 8.5 8.6 8.7 8.8 Carrano, Matthew T.; Sampson, Scott D. The Phylogeny of Ceratosauria (Dinosauria: Theropoda). Journal of Systematic Palaeontology. January 2008, 6 (2): 183–236. doi:10.1017/S1477201907002246.
- ^ 9.0 9.1 9.2 Paul, Gregory S. Predatory Dinosaurs of the World. 1988: 284–285. ISBN 978-0-671-61946-6.
- ^ Yong, Ed. Butch tail made Carnotaurus a champion dinosaur sprinter. National Geographic. October 18, 2011 [July 31, 2019]. (原始內容存檔於2021-01-23). (頁面存檔備份,存於網際網路檔案館)
- ^ Headden, Jaime A. Re: Carnotaurus sastrei etymology. Dinosaur Mailing List. September 19, 2006 [2021-01-31]. (原始內容存檔於2012-02-22). (頁面存檔備份,存於網際網路檔案館)
- ^ Bonaparte, José F. The gondwanian theropod families Abelisauridae and Noasauridae. Historical Biology. 1991, 5: 1. doi:10.1080/10292389109380385.
- ^ 13.0 13.1 Bonaparte, José F. Cretaceous tetrapods of Argentina. Münchener Geowissenschaftliche Abhandlung. 1996, A (30): 89.
- ^ Glut, Donald F. Carnotaurus. Dinosaurs, the encyclopedia. Jefferson, North Carolina: McFarland & Company, Inc. Publishers. 1997: 256–259. ISBN 978-0-375-82419-7.
- ^ 15.0 15.1 Glut, Donald F. Carnotaurus. Dinosaurs: The Encyclopedia. 3rd Supplement. Jefferson, North Carolina: McFarland & Company, Inc. 2003: 274–276. ISBN 978-0-7864-1166-5.
- ^ Glut, Donald F. Carnotaurus. Dinosaurs: The Encyclopedia. 1st Supplement. Jefferson, North Carolina: McFarland & Company, Inc. 2000: 165–167. ISBN 978-0-7864-0591-6.
- ^ 17.0 17.1 Candeiro, Carlos Roberto dos Anjos; Martinelli, Agustín Guillermo. Abelisauroidea and carchardontosauridae (theropoda, dinosauria) in the cretaceous of south america. Paleogeographical and geocronological implications. Uberlândia: 5–19.
- ^ 18.0 18.1 18.2 Juárez Valieri, Rubén D.; Porfiri, Juan D.; Calvo, Jorge O. New information on Ekrixinatosaurus novasi Calvo et al. 2004, a giant and massively-constructed Abelisauroid from the "Middle Cretaceous" of Patagonia. Paleontologıa y Dinosaurios en América Latina. 2010: 161–169.
- ^ 19.0 19.1 19.2 Paul, Gregory S. The Princeton Field Guide to Dinosaurs 1st. Princeton University Press. 2010. ISBN 9780691137209.
- ^ Calvo, Jorge O.; Rubilar-Rogers, David; Moreno, Karen. A new Abelisauridae (Dinosauria: Theropoda) from northwest Patagonia (PDF). Ameghiniana. 2004, 41 (4): 555–563. (原始內容 (PDF)存檔於March 9, 2012). (頁面存檔備份,存於網際網路檔案館)
- ^ Grillo, O.N.; Delcourt, R. Allometry and body length of abelisauroid theropods: Pycnonemosaurus nevesi is the new king. Cretaceous Research. 2016, 69: 71–89. doi:10.1016/j.cretres.2016.09.001.
- ^ 22.0 22.1 (英文)Mazzetta, G.V., Christiansen, P.& Farin, R.;Giants and Bizarres:Body Size of Some Southern South American Cretaceous Dinosaurs (頁面存檔備份,存於網際網路檔案館); Historical Biology, 2004, 1–13
- ^ 23.0 23.1 23.2 23.3 23.4 23.5 23.6 23.7 Mazzetta, Gerardo V.; Fariña, Richard A.; Vizcaíno, Sergio F. On the palaeobiology of the South American horned theropod Carnotaurus sastrei Bonaparte (PDF). Gaia. 1998, 15: 185–192 [2017-03-04]. (原始內容存檔 (PDF)於2015-09-23). (頁面存檔備份,存於網際網路檔案館)
- ^ 24.0 24.1 24.2 Mazzetta, Gerardo V.; Christiansen, Per; Fariña, Richard A. Giants and Bizarres: Body size of some southern South American Cretaceous dinosaurs (PDF). Historical Biology. 2004, 16 (2): 71–83 [2008-06-15]. doi:10.1080/08912960410001715132. (原始內容存檔 (PDF)於2018-08-19). (頁面存檔備份,存於網際網路檔案館)
- ^ 25.0 25.1 25.2 25.3 Cerroni, Mauricio A.; Paulina-Carabajal, Ariana. Novel information on the endocranial morphology of the abelisaurid theropod Carnotaurus sastrei. Comptes Rendus Palevol. 2019, 18 (8): 985–995. doi:10.1016/j.crpv.2019.09.005.
- ^ 26.0 26.1 Sampson, Scott D.; Witmer, Lawrence M. Craniofacial Anatomy of Majungasaurus crenatissimus (Theropoda: Abelisauridae) From the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology. 2007, 27 (sp8): 95–96. doi:10.1671/0272-4634(2007)27[32:CAOMCT]2.0.CO;2.
- ^ 27.0 27.1 27.2 Hartman, Scott. Carnotaurus – delving into self-parody?. 2012 [December 7, 2012]. (原始內容存檔於2013-06-03). (頁面存檔備份,存於網際網路檔案館)
- ^ Méndez, Ariel. The cervical vertebrae of the Late Cretaceous abelisaurid dinosaur Carnotaurus sastrei (PDF). Acta Palaeontologica Polonica. 2014, 59 (1): 99–107 [December 30, 2012]. doi:10.4202/app.2011.0129. (原始內容 (PDF)存檔於April 3, 2013). (頁面存檔備份,存於網際網路檔案館)
- ^ 29.0 29.1 29.2 29.3 29.4 Persons, W.S.; Currie, P.J. Farke, Andrew Allen , 編. Dinosaur Speed Demon: The caudal musculature of Carnotaurus sastrei and implications for the evolution of South American abelisaurids. PLOS ONE. 2011, 6 (10): e25763. Bibcode:2011PLoSO...625763P. PMC 3197156
 . PMID 22043292. doi:10.1371/journal.pone.0025763.
. PMID 22043292. doi:10.1371/journal.pone.0025763.
- ^ 30.0 30.1 30.2 Ruiz, Javier; Torices, Angélica; Serrano, Humberto; López, Valle. The hand structure of Carnotaurus sastrei (Theropoda, Abelisauridae): implications for hand diversity and evolution in abelisaurids (PDF). Palaeontology. 2011, 54 (6): 1271–1277 [2021-01-31]. doi:10.1111/j.1475-4983.2011.01091.x. (原始內容存檔 (PDF)於2017-09-22). (頁面存檔備份,存於網際網路檔案館)
- ^ Agnolin, Federico L.; Chiarelli, Pablo. The position of the claws in Noasauridae (Dinosauria: Abelisauroidea) and its implications for abelisauroid manus evolution. Paläontologische Zeitschrift. June 2010, 84 (2): 293–300. doi:10.1007/s12542-009-0044-2.
- ^ Senter, P. Vestigial skeletal structures in dinosaurs. Journal of Zoology. 2010, 280 (4): 60–71. doi:10.1111/j.1469-7998.2009.00640.x.
- ^ Kevin M. Middleton; Stephen M. Gatesy. Theropod forelimb design and evolution. Zoological Journal of the Linnean Society (The Linnean Society of London). Feb 2000, 128 (2): 149–187 [2017-03-04]. doi:10.1111/j.1096-3642.2000.tb00160.x. (原始內容存檔於2017-03-11). (頁面存檔備份,存於網際網路檔案館)(英文)
- ^ (英文)http://dml.cmnh.org/2001Apr/msg00486.html (頁面存檔備份,存於網際網路檔案館) Post de Archives of the DINOSAUR Mailing List sobre la posición de los brazos del Carnotaurus
- ^ 35.0 35.1 35.2 35.3 Canale, Juan I.; Scanferla, Carlos A.; Agnolin, Federico; Novas, Fernando E. New carnivorous dinosaur from the Late Cretaceous of NW Patagonia and the evolution of abelisaurid theropods. Naturwissenschaften. 2009, 96 (3): 409–14. Bibcode:2009NW.....96..409C. PMID 19057888. doi:10.1007/s00114-008-0487-4. hdl:11336/52024.
- ^ 36.0 36.1 Coria, Rodolfo A.; Chiappe, Luis M.; Dingus, Lowell. A new close relative of Carnotaurus sastrei Bonaparte 1985 (Theropoda: Abelisauridae) from the Late Cretaceous of Patagonia. Journal of Vertebrate Paleontology. 2002, 22 (2): 460. doi:10.1671/0272-4634(2002)022[0460:ANCROC]2.0.CO;2.
- ^ Ezcurra, Martín D.; Agnolin, Federico L.; Novas, Fernando E. An abelisauroid dinosaur with a non-atrophied manus from the Late Cretaceous Pari Aike Formation of southern Patagonia (PDF). Zootaxa. 2010, 2450: 14 [2021-01-31]. doi:10.11646/zootaxa.2450.1.1. (原始內容存檔 (PDF)於2012-09-19). (頁面存檔備份,存於網際網路檔案館)
- ^ 38.0 38.1 38.2 Delcourt, Rafael. Ceratosaur palaeobiology: new insights on evolution and ecology of the southern rulers. Scientific Reports. 2018, 8 (1): 9730. Bibcode:2018NatSR...8.9730D. PMC 6021374 . PMID 29950661. doi:10.1038/s41598-018-28154-x.
- ^ 39.0 39.1 Sereno, Paul C.; Wilson, Jeffrey A.; Conrad, Jack L. New dinosaurs link southern landmasses in the Mid-Cretaceous. Proceedings of the Royal Society B: Biological Sciences. July 7, 2004, 271 (1546): 1325–1330. PMC 1691741 . PMID 15306329. doi:10.1098/rspb.2004.2692.
- ^ Tykoski, Ronald B.; Rowe, Timothy. Ceratosauria. Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (編). The Dinosauria Second. Berkeley: University of California Press. 2004: 65. ISBN 978-0-520-24209-8.
- ^ Wilson, Jeffrey A.; Sereno, Paul C.; Srivastava, Suresh; Bhatt, Devendra K.; Khosla, Ashu; Sahni, Ashok. A new abelisaurid (Dinosauria, Theropoda) from the Lameta Formation (Cretaceous, Maastrichtian) of India. Contributions from the Museum of Paleontology. 2003, 31 (1): 25. hdl:2027.42/48667.
- ^ Sereno, Paul C. A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria. Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 10 November 1998, 210 (1): 41–83. doi:10.1127/njgpa/210/1998/41.
- ^ Sereno, Paul. Carnotaurinae. Taxon Search. 2005 [December 29, 2012]. (原始內容存檔於May 16, 2012). (頁面存檔備份,存於網際網路檔案館)
- ^ Sereno, Paul. Carnotaurini. Taxon Search. 2005 [December 29, 2012]. (原始內容存檔於May 16, 2012). (頁面存檔備份,存於網際網路檔案館)
- ^ Wang, S.; Stiegler, J.; Amiot, R.; Wang, X.; Du, G.-H.; Clark, J.M.; Xu, X. Extreme ontogenetic changes in a ceratosaurian theropod (PDF). Current Biology. 2017, 27 (1): 144–148. PMID 28017609. doi:10.1016/j.cub.2016.10.043.
- ^ Tortosa, Thierry; Eric Buffetaut; Nicolas Vialle; Yves Dutour; Eric Turini; Gilles Cheylan. A new abelisaurid dinosaur from the Late Cretaceous of southern France: Palaeobiogeographical implications. Annales de Paléontologie. 2013, 100 (In press): 63–86. doi:10.1016/j.annpal.2013.10.003.
- ^ Filippi, Leonardo S.; Méndez, Ariel H.; Juárez Valieri, Rubén D.; Garrido, Alberto C. A new brachyrostran with hypertrophied axial structures reveals an unexpected radiation of latest Cretaceous abelisaurids (PDF). Cretaceous Research. 2016-06-01, 61: 209–219 [2021-01-31]. ISSN 0195-6671. doi:10.1016/j.cretres.2015.12.018. (原始內容存檔 (PDF)於2021-01-09) (英語). (頁面存檔備份,存於網際網路檔案館)
- ^ 48.0 48.1 48.2 48.3 48.4 48.5 48.6 Mazzetta, Gerardo V.; Cisilino, Adrián P.; Blanco, R. Ernesto; Calvo, Néstor. Cranial mechanics and functional interpretation of the horned carnivorous dinosaur Carnotaurus sastrei. Journal of Vertebrate Paleontology. 2009, 29 (3): 822–830. doi:10.1671/039.029.0313.
- ^ Chure, Daniel J. On the orbit of theropod dinosaurs. Gaia. 1998, 15: 233–240.
- ^ Snively, Eric; Cotton, John R.; Witmer, Lawrence; Ridgely, Ryan; Theodor, Jessica. Finite element comparison of cranial sinus function in the dinosaur Majungasaurus and head-clubbing giraffes. Summer Bioengineering Conference 54587. American Society of Mechanical Engineers: 1075–1076. 2011.
- ^ Rayfield, E. J., Norman, D. B., Horner, C. C., Horner, J. R., Smith, P. M., Thomason, J. J. and Upchurch, P. 2001. Cranial design and function in a large theropod dinosaur. Nature 409: 1033-1037.
- ^ (英文)Barret, Paul M. & Rayfield, Emily J.; Ecological and evolutionary implications of dinosaur feeding behaviour; TRENDS in Ecology and Evolution Vol.21 No.4 April 2006
- ^ Therrien, François; Henderson, Donald; Ruff, Christopher. Bite Me – Biomechanical Models of Theropod Mandibles and Implications for Feeding Behavior. Carpenter, Kenneth (編). The carnivorous dinosaurs. Indiana University Press. 2005: 179–198, 228. ISBN 978-0-253-34539-4.
- ^ Bakker, Robert T. Brontosaur killers: Late Jurassic allosaurids as sabre-tooth cat analogues (PDF). Gaia. 1998, 15: 145–158 [2021-01-31]. (原始內容存檔 (PDF)於2021-04-24). (頁面存檔備份,存於網際網路檔案館)
- ^ Mazzetta, Gerardo V.; Farina, Richard A. Estimacion de la capacidad atlética de Amargasaurus cazaui Salgado y Bonaparte, 1991, y Carnotaurus sastrei Bonaparte, 1985 (Saurischia, Sauropoda-Theropoda). XIV Jornadas Argentinas de Paleontologia de Vertebrados, Ameghiniana. 1999, 36 (1): 105–106 (西班牙語).
- ^ Predatory dinosaur was fearsomely fast. CBC News. October 21, 2011 [April 22, 2017]. (原始內容存檔於2020-12-22). (頁面存檔備份,存於網際網路檔案館)
- ^ Matthew C. Lamanna; Rubén D. Martínez and Joshua B. Smith. A Definitive Abelisaurid Theropod Dinosaur from the Early Late Cretaceous of Patagonia. Journal of Vertebrate Paleontology (The Society of Vertebrate Paleontology). Mar 14, 2002, 22 (1): 58–69 [2017-03-04]. (原始內容存檔於2020-07-04). (頁面存檔備份,存於網際網路檔案館)(英文)(西班牙文)
- ^ Le Loeuff, Jean. Biogeography. Padian, Kevin; Currie, Philip J. (編). Encyclopedia of dinosaurs. San Diego: Academic Press. 1997: 51–56. ISBN 978-0-12-226810-6.
- ^ 59.0 59.1 59.2 Pascual, Rosendo; Goin, Francisco J.; González, Pablo; Ardolino, Alberto; Puerta, Pablo F. A highly derived docodont from the Patagonian Late Cretaceous: evolutionary implications for Gondwanan mammals. Geodiversitas. 2000, 22 (3): 395–414 [2021-01-31]. (原始內容存檔於2021-03-02). (頁面存檔備份,存於網際網路檔案館)
- ^ Sterli, Juliana; De la Fuente, Marcelo S. A new turtle from the La Colonia Formation (Campanian–Maastrichtian), Patagonia, Argentina, with remarks on the evolution of the vertebral column in turtles. Palaeontology. 2011, 54 (1): 65. doi:10.1111/j.1475-4983.2010.01002.x.
- ^ Albino, Adriana M. New record of snakes from the Cretaceous of Patagonia (Argentina). Geodiversitas. 2000, 22 (2): 247–253 [2021-01-31]. (原始內容存檔於2021-02-14). (頁面存檔備份,存於網際網路檔案館)
- ^ 62.0 62.1 Gasparini, Zulma; De la Fuente, Marcelo. Tortugas y Plesiosaurios de la Formación La Colonia (Cretácico Superior) de Patagonia, Argentina. Revista Española de Paleontología. 2000, 15 (1): 23 (西班牙語).
- ^ Kielan−Jaworowska, Zofia; Ortiz−Jaureguizar, Edgardo; Vieytes, Carolina; Pascual, Rosendo; Goin, Francisco J. First ?cimolodontan multi−tuberculate mammal from South America (PDF). Acta Palaeontologica Polonica. 2007, 52 (2): 257–262 [2021-01-31]. (原始內容存檔 (PDF)於2017-08-08). (頁面存檔備份,存於網際網路檔案館)
- ^ Lawver, Daniel R.; Debee, Aj M.; Clarke, Julia A.; Rougier, Guillermo W. A New Enantiornithine Bird from the Upper Cretaceous La Colonia Formation of Patagonia, Argentina. Annals of Carnegie Museum. January 1, 2011, 80 (1): 35–42. doi:10.2992/007.080.0104.
- ^ (英文)Gandolfo Maria A.,Ruben; N.; Flora of the La Colonia Formation (Maastrichtian, Late Cretaceus), Chubut, Patagonia, Argentina
- ^ (西班牙文)Prámparo, M.B., Quattrocchio, M, Gandolfo, M.A., Zamaloa M. Del C., Y Romero, E.; Historia evolutiva de las angiospermas (Cretácico-Paleógeno) en Argentina a través de los registros paleoflorísticos; Asociación Paleontológica Argentina. Publicación Especial 11; Ameghiniana 50º aniversario: 157-172. Buenos Aires, 25-11-2007; ISSN 0328-347X
- ^ (英文) Archangelsky, Ana, Phipps, Carlie J, Taylor, Thomas N &, Edith L.;Paleoazolla, a new heterosporous fern from the Upper Cretaceous of Argentina[永久失效連結]; American Journal of Botany. 1999;86:1200-1206.
- ^ (英文)Gandolfo Maria A. & Cuneo Ruben N.; Fossil Nelumbonaceae from the La Colonia Formation (Campanian–Maastrichtian, Upper Cretaceous), Chubut, Patagonia, Argentina; Review of Palaeobotany and Palynology, Volume 133, Issues 3-4, February 2005, Pages 169-178
- ^ (英文)Crichton, Michael. The Lost World. Ballantine Books. September 1995: 416 pp. ISBN 0-679-41946-2.
- ^ 王誦賢. 【侏羅紀世界2】導演揚言今集恐龍品種最多 中國角龍都有份登場. 香港01. 2018-06-03 [2019-07-20]. (原始內容存檔於2020-10-29). (頁面存檔備份,存於網際網路檔案館)
- ^ (英文)Eric Leighton & Ralph Zondag(Director). Dinosaur (motion picture). USA: Walt Disney Pictures. 2000-05-19 [2008-02-03]. (原始內容存檔於2021-05-04).
外部連結
[編輯]- (西班牙文)MEF - 食肉牛龍的特徵
- (英文)食肉牛龍的簡介與想像圖
- (英文)食肉牛龍 - 獸腳亞目資料庫
- (英文)食肉牛龍的簡介
- (英文)食肉牛龍 - DinoData (頁面存檔備份,存於網際網路檔案館)
- (英文)阿貝力龍科 - The Thescelsosaurus
- (英文)各種恐龍的學名與發音
- (西班牙文)馬德普拉塔國立大學的食肉牛龍咬合力研究
- (英文)史考特·哈特曼(Scott Hartman)所製作的骨骼重建圖 (頁面存檔備份,存於網際網路檔案館)

.jpg)













{kind=link}
{kind=link}